Torque, \( \tau \) (Greek symbol tau), also called moment of force, is a physical quantity that measures the rotational effect of a force applied to an object around a pivot point or axis. It is the rotational equivalent of linear force, causing an object to rotate or twist rather than move in a straight line.
Torque Formula |
||
|
\( \tau \;=\; r \cdot F \cdot sin( \theta ) \) (Torque) \( r \;=\; \dfrac{ \tau }{ F \cdot sin ( \theta ) } \) \( F \;=\; \dfrac{ \tau }{ r \cdot sin ( \theta ) } \) \( \theta \;=\; \dfrac{ \tau }{ r \cdot F } \) |
||
| Symbol | English | Metric |
| \( \tau \) (Greek symbol tau) = Torque | \(lbf-ft\) | \(N-m\) |
| \( r \) = Radius is the Distance from the Pivot Point | \(in\) | \(mm\) |
| \( F \) = Magnitude of the Force Applied | \(lbf\) | \(N\) |
| \( \theta \) (Greek symbol theta) = Angle Between the Force Vector and the Leverage Arm Vector | \(deg\) | \(rad\) |
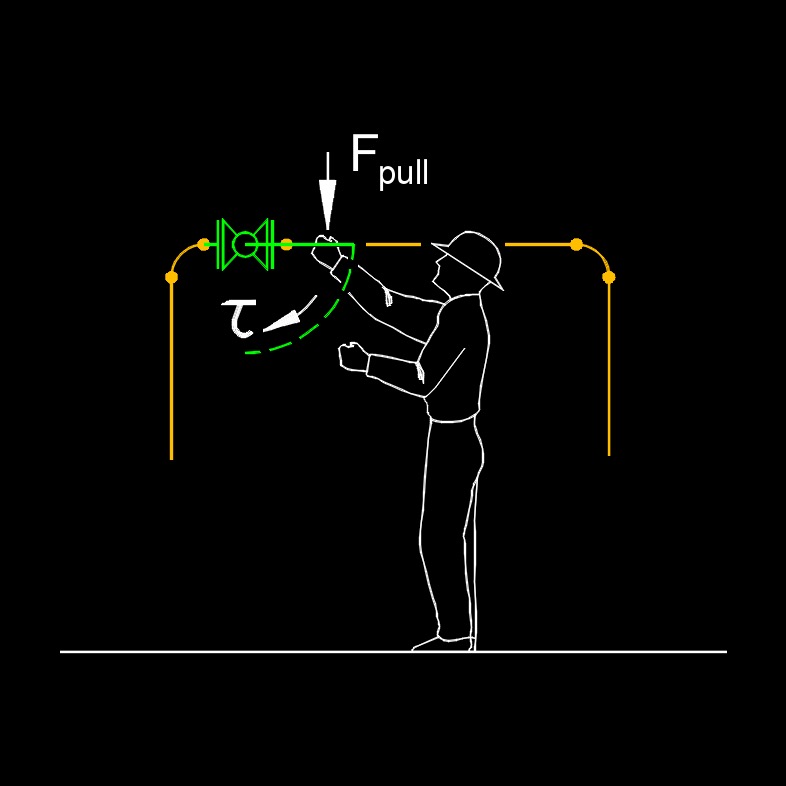 Torque depends on two key factors: the magnitude of the force applied and the distance from the axis of rotation to the point where the force is applied, known as the lever arm or moment arm. Torque plays a critical role in many applications, such as in engines, where it quantifies the turning power that drives machinery, or in everyday tasks like tightening a bolt with a wrench, where a longer wrench increases the lever arm, making it easier to apply torque. Essentially, torque determines how effectively a force can cause or change rotational motion, making it a fundamental concept in mechanics and engineering.
Torque depends on two key factors: the magnitude of the force applied and the distance from the axis of rotation to the point where the force is applied, known as the lever arm or moment arm. Torque plays a critical role in many applications, such as in engines, where it quantifies the turning power that drives machinery, or in everyday tasks like tightening a bolt with a wrench, where a longer wrench increases the lever arm, making it easier to apply torque. Essentially, torque determines how effectively a force can cause or change rotational motion, making it a fundamental concept in mechanics and engineering.
Torque Examples
- A force applied at the handle of a wrench produces torque about the center of the bolt. Increasing the wrench length increases the torque for the same applied force.
- Rotating the valve wheel or lever applies torque to the valve stem, allowing the valve to open or close.
- An electric, hydraulic, pneumatic, or internal-combustion motor produces torque on a rotating shaft to perform mechanical work.
- Combustion forces acting through connecting rods generate torque on the crankshaft, which ultimately drives the vehicle.
- Torque is transmitted and often modified in magnitude through gears to drive machinery or vehicles.
- The engine supplies torque to the rotor system, causing the rotor blades to rotate and generate lift.
- Forces acting at distances from the pivot create torque about the fulcrum.
- Gravity produces a torque about the pivot point, causing the pendulum to swing toward equilibrium.
- Forces from loads and counterweights create torques about the boom's pivot point.
![]()